
Тасманијски ђаво
| Tasmanijski đavo Vremenski raspon: kasni holocen
| |
|---|---|

| |
| mužjak | |

| |
| Tasmanijski đavo | |
| Naučna klasifikacija | |
| Domen: | Eukaryota |
| Carstvo: | Animalia |
| Tip: | Chordata |
| Klasa: | Mammalia |
| Infraklasa: | Marsupialia |
| Red: | Dasyuromorphia |
| Porodica: | Dasyuridae |
| Rod: | Sarcophilus |
| Vrsta: | S. harrisii
|
| Binomno ime | |
| Sarcophilus harrisii | |

| |
| Distribucija tasmanijskog đavola (sivo) | |
Tasmanijski đavo (lat. Sarcophilus harrisii) je vrsta torbara mesoždera sa Tasmanije. Jedini je živeći predstavnik roda Sarcophilus. Posle izumiranja tilacina, postao je najveći torbar mesožder u svetu. Ima crno krzno, a poznat je po glasnim i zastrašujućim kricima koje ispušta. Iz Australije je nestao pre više od 3 000 godina opstavši samo na Tasmaniji. Zbog pretnje stoci, bio je lovljen sve do 1941. godine, kada je stavljen pod zaštitu. Od kasnih 1990-ih, među tasmanijskim đavolima je sve zastupljeniji facijalni tumor, zbog čega je ova vrsta u opasnosti od izumiranja.
Taksonomija
[uredi | uredi izvor]Verujući da je to vrsta oposuma, prirodnjak Džordž Haris napisao je prvi objavljeni opis tasmanskog đavola 1807. godine, nazvavši ga Didelphis ursina,[3] zbog njegovih karakteristika poput medveda, kao što je okruglo uvo.[4] Ranije je održao prezentaciju na ovu temu u Zoološkom društvu u Londonu.[5] Međutim, Džordž Šo je 1800. dao to posebno binomsko ime običnom vombatu (kasnije preklasifikovanom kao Vombatus ursinus), i stoga nije bilo dostupno.[6] Godine 1838, Ričard Oven je primerak nazvao Dasyurus laniarius,[7] ali ga je do 1877. preneo u Sarcophilus. Savremenog tasmanijskog đavola nazvao je Sarcophilus harrisii („Harisov ljubitelj mesa“) od strane francuskog prirodnjaka Pjera Boitara 1841. godine.[8]
Kasnija revizija đavolje taksonomije, objavljena 1987. godine, pokušala je da promeni ime vrste u Sarcophilus laniarius na osnovu kopnenih fosilnih zapisa samo nekoliko životinja.[9] Međutim, to nije prihvaćeno od strane taksonomske zajednice u celini; naziv S. harrisii je zadržan, a S. laniarius upotrebljeno za fosilnu vrstu.[6] „Vilzebubovo štene” je bilo rano narodno ime koje su mu dali istraživači Tasmanije, u vezi sa religioznom figurom koja je princ pakla i pomoćnik Satane;[5] istraživači su prvi put naišli na životinju čuvši njenu daleko-dostižući vokalizaciju noću.[10] Srodna imena koja su korišćena u 19. veku bila su Sarcophilus satanicus („satanistički ljubitelj mesa“) i Diabolus ursinus („medveđi đavo“), a sve zbog ranih pogrešnih shvatanja ove vrste kao neumoljivo opake.[5] Tasmanijski đavo (Sarcophilus harrisii) pripada porodici Dasyuridae. Rod Sarcophilus sadrži još dve vrste, poznate samo iz pleistocenskih fosila: S. laniarius i S. moomaensis. Filogenetska analiza pokazuje da je tasmanski đavo najbliži srodnik kvola.[11]
Prema Pembertonu, mogućim precima đavola je možda bilo potrebno da se penju na drveće da bi došli do hrane, što je dovelo do povećanja veličine i skakajućeg hoda mnogih torbara. On je spekulisao da su ove adaptacije možda izazvale neobičan hod savremenog đavola.[12] Teoretiše se da se specifična loza tasmanskog đavola pojavila tokom miocena, pri čemu molekularni dokazi ukazuju na odvajanje od predaka kvola pre između 10 i 15 miliona godina,[13] kada su se ozbiljne klimatske promene odvijale u Australiji, transformišući klimu od toplog i vlažnog do sušnog, suvog ledenog doba, što je rezultiralo masovnim izumiranjem.[12] Pošto je većina njihovog plena umrla od hladnoće, preživelo je samo nekoliko mesoždera, uključujući pretke kvola i torbarskog vuka. Nagađa se da je đavolja loza možda nastala u ovom trenutku kako bi popunila nišu u ekosistemu, kao čistač koji je uklanjao strvinu koju bi iza sebe ostavio torbarski vuk koji selektivno jede.[12] Izumrli Glaucodon ballaratensis iz pliocenskog doba nazvan je posrednom vrstom između kvola i đavola.[14]

Fosilne naslage u krečnjačkim pećinama u Narakurtu, Južna Australija, koje datiraju iz miocena uključuju primerke S. laniarius, koji su bili oko 15% veći i 50% teži od savremenih đavola.[15] Stariji primerci za koje se veruje da su stari 50–70.000 godina pronađeni su u Darling Daunsu u Kvinslendu i u Zapadnoj Australiji.[16] Nije jasno da li je moderni đavo evoluirao od S. laniarius, ili su oni u to vreme koegzistirali.[16] Ričard Oven je zastupao potonju hipotezu u 19. veku, na osnovu fosila pronađenih 1877. u Novom Južnom Velsu.[16] Velike kosti koje se pripisuju S. moornaensis pronađene su u Novom Južnom Velsu,[16] i pretpostavlja se da su ove dve izumrle veće vrste možda lovile i sakupljale leševe.[16] Poznato je da je pre milionima godina postojalo nekoliko rodova torbarskog vuka i da su bili različiti po veličini, a manji su se više oslanjali na traženje hrane.[17] Pošto su đavo i torbarski vuk slični, izumiranje koegzistirajućih rodova torbarskog vuka je citirano kao dokaz za analognu istoriju đavola.[18] Spekulisalo se da im je manja veličina S. laniarius i S. moornaensis omogućila da se efikasnije prilagode promenljivim uslovima i da prežive duže od odgovarajućih torbarskih vukova.[18] Pošto je izumiranje ove dve vrste došlo u isto vreme kada su ljudi živeli u Australiji, lov od strane ljudi i čišćenje zemljišta su razmatrani kao mogući uzroci.[19] Kritičari ove teorije ističu da su autohtoni Australijanci razvili bumerange i koplja za lov samo pre oko 10.000 godina, da je malo verovatan kritičan pad broja usled sistematskog lova. Oni takođe ističu da pećine koje naseljavaju Aboridžini imaju nizak udeo kostiju i slika đavola na stenama, i sugerišu da je to pokazatelj da to nije bio veliki deo starosedeoca života. Jedan naučni izveštaj iz 1910. tvrdio je da Aboridžini više vole meso biljojeda nego mesoždera.[20] Druga zastupljena teorija za izumiranje je bila da je to bilo zbog klimatskih promena koje je izazvalo najnedavnije ledeno doba.[19]
Genetika
[uredi | uredi izvor].png)
Genom tasmanskog đavola je sekvencioniran 2010. godine od strane Velkom Trast Sanger instituta.[21] Kao i svi dasjuridi, đavo ima 14 hromozoma.[22] Đavoli imaju nizak genetski diverzitet u poređenju sa drugim australijskim torbarima i placentnim mesožderima; ovo je u skladu sa efektom osnivača jer su rasponi veličine alela bili niski i skoro kontinuirani kroz sve merene podpopulacije. Alelni diverzitet je izmeren na 2,7–3,3 u uzorkovanim podpopulacijama, a heterozigotnost je bila u opsegu 0,386–0,467.[23] Prema studiji Mena Džounsa, „protok gena izgleda opsežan do 50 km (31 mi)”, što znači visoku stopu dodeljivanja izvornim ili bliskim susednim populacijama „u skladu sa podacima o kretanju. U većim razmerama (150–250 km ili 93–155 mi), protok gena je smanjen, ali nema dokaza za izolaciju usled udaljenosti”.[23] Ostrvski efekti su takođe možda doprineli njihovoj niskoj genetskoj raznovrsnosti. Periodi niske gustine naseljenosti su takođe mogli stvoriti umerena uska grla u populaciji, smanjujući genetski diverzitet.[23] Smatra se da je nizak genetski diverzitet karakterističan za populaciju tasmanskog đavola još od sredine holocena.[24] Epidemije đavoljeg tumora lica (DFTD) izazivaju povećanje inbridinga.[25] Podpopulacija đavola na severozapadu države genetski se razlikuje od drugih đavola,[26] ali postoji izvesna razmena između dve grupe.[27]
Analiza polimorfizma konformacije lanca (OSCP) na domenu klase I glavnog kompleksa histokompatibilnosti (MHC), uzeta sa različitih lokacija širom Tasmanije, pokazala je 25 različitih tipova i pokazala različit obrazac MHC tipova u severozapadnoj Tasmaniji do istočne Tasmanije. Đavoli na istoku države imaju manje MHC raznolikosti; 30% je istog tipa kao tumor (tip 1), a 24% je tipa A.[28] Sedam od svakih deset đavola na istoku su tipa A, D, G ili 1, koji su povezani sa DFTD; dok samo 55% zapadnih đavola spada u ove MHC kategorije. Od 25 MHC tipova, 40% je isključivo za zapadne đavole. Iako je severozapadna populacija generalno manje genetski raznolika, ona ima veću raznolikost MHC gena, što im omogućava da podignu imuni odgovor na DFTD. Prema ovom istraživanju, mešanje đavola može povećati šanse za bolest.[28] Od petnaest različitih regiona na Tasmaniji obuhvaćenih ovim istraživanjem, šest je bilo u istočnoj polovini ostrva. U istočnoj polovini, Eping šuma je imala samo dva različita tipa, od kojih je 75% bilo tip O. U oblasti Bakland-Nagent, bila su prisutna samo tri tipa, a bilo je u proseku 5,33 različita tipa po lokaciji. Nasuprot tome, na zapadu, Kejp Korel je dao tri vrste, a Togari Nort-Kristmas Hils je dao šest, ali ostalih sedam lokacija imalo je najmanje osam MHC tipova, a Vest Pensil Pajn je imao 15 tipova. U proseku je bilo 10,11 tipova MHC po lokaciji na zapadu.[28] Nedavna istraživanja su pokazala da divlja populacija đavola brzo razvija otpor prema DFTD.[29]
Fizički opis
[uredi | uredi izvor]
Tasmanski đavo je najveći živi tobolčar-mesojed u Australiji. Zdepaste je građe, sa velikom glavom i repom koji čini polovinu dužine tela. Neobično za torbara, prednji udovi su nešto duži od zadnjih. Mogu trčati brzinom do 13 km na sat na kratke razdaljine. Krzno je obično crno, a nepravilne bele šare na prsima i trtici su česte. Mužjaci su obično veći od ženki, sa prosečnom dužinom od 652 mm od glave do početka repa. Rep je dug 258 mm. Prosečna težina je 8 kg. Prosečna dužina tela i glave kod ženke je 570 mm, sa repom od 244 mm i prosečnom težinom od 6 kg.[30] Prosečna dužina života tasmanskog đavola u divljini je 6 godina, ali u zatočeništvu duže žive.
Tasmanski đavo nakuplja masti u svom repu, pa nezdravi đavoli obično imaju tanke repove. Koristi mirisne žlezde pri bazi repa da označe tlo iza sebe mirisom. Kada ga se uznemirava, đavo ispusti odvratan smrad, skoro isto tako smrdljiv kao kod tvora.
Đavo ima duge brkove na licu i grude na glavi. Pomažu mu da nađe plen u mraku, a pokazuju i koliko je drugi đavo blizu dok se hrane. Sluh je dominantno čulo, a takođe ima i odlično čulo mirisa. Kako tasmanski đavoli love noću, izgleda da vide crno-belo. Tako mogu lahko videti pokrete, ali imaju teškoća kada gledaju predmete koji se ne pomeraju.[31]
Jedna analiza snage ugriza kod sisara u odnosu na veličinu tela pokazuje da đavo ima najjači ugriz, preko 35.000 kPa.[32] Snaga ugriza je tako velika jer je i glava velika. Takođe ima i zube koji rastu polako tokom života. Zubi i vilice tasmanskog đavola i hijene su slični, što je jedan primer konvergentne evolucije.[33]
Ishrana
[uredi | uredi izvor]Love svoj plen sami, ali jedu u grupi, pritom prave veliku buku, ispuštaju hrapave glasove koji se čuju kilometrima daleko, jer se oko lovine okupi i do 12 jedinki. Nisu probirljivi što se izbora hrane tiče, love i jedu vombate, male sisare, gmizavce, ribe, žabe, insekte i čak domaće životinje, npr. leševe ovaca. Jedu mnogo, pojedu u proseku hrane količinski u vrednosti od 15% njihove ukupne telesne mase, a mogu pojesti i do 40% svoje ukupne mase. Vrlo su im male potrebe za vodom. Spadaju u životinje sa najmanjom potrošnjom vode, tj. sa najmanjom potrebom.
Stanište
[uredi | uredi izvor]Naseljavaju čitavu teritoriju Tasmanije, ima ih i u urbanim naseljima. Posebno im pogoduju suve šumske oblasti i obalna, šumom okružena područja.
Rak lica
[uredi | uredi izvor]Godine 1995. tumor lica tasmanijskog đavola opustošio je populaciju ovih životinja u Tasmaniji. Oko 65% populacije je bilo zaraženo. Smrtnost je 100% u populaciji od 12 - 18 meseci života. Poreklo ove vrste raka je još uvek nepoznato, istraživanja se sprovode, monitoring vrste postoji. Iz populacija se odstranjuju sve bolesne jedinke, radi sprečavanja širenja zaraze, jer je utvrđeno da je bolest „prelazna“. Životinje se zaraze kontaktom ili tokom zajedničke ishrane. Do smrti dolazi jer bolest oštećuje u velikoj meri njihovo lice, kada više ne mogu da se hrane, umiru od gladi. Jedan od uzroka može biti i izuzetno niska stopa genetičke raznovrsnosti i hromozomske mutacije jedinstvene i češće među karnivornim sisarima. Potpuno zdrave jedinke izdvojene su i gaje se zaštićene i nadzirane od strane Australijske vlade, za slučaj da dođe do izumiranja populacije koja živi slobodno u prirodi.
Ponašanje
[uredi | uredi izvor]Aktivnost i teritorijalnost
[uredi | uredi izvor]Aktivni su u sumrak i tokom noći, tada love svoj plen. Tokom dana se odmaraju i dremaju u žbunju ili jazbinama. Teritorijalne su životinje, zauzimaju prostor od 8 - 20 km², ali se on može preklapati i sa teritorijom drugih životinja. Obeležavaju svoju teritoriju svojim urinom ili izmetom, daju do znanja drugim jedinkama da je određena jazbina zauzeta.
Socijalna struktura
[uredi | uredi izvor]Solitarne su životinje, grupišu se samo tokom ishrane, jedinke zajedno komadaju plen.
Hijerarhija
[uredi | uredi izvor]Obično ustanovljuju dominaciju svojim stavom i glasnim zvukovima, gde dolazi i do međusobnih okršaja, koji su česti u
sezoni parenja jednako kao i za vreme ishrane.
Zvučna i vizuelna komunikacija
[uredi | uredi izvor]Sluh je kod njih izuzetno dobro razvijen. Proučavanjem ovih životinja tokom hranjenja, primećeno je 20-ak različitih položaja tela, među kojima je i karakteristično zlobno, jogunasto zevanje i 11 različitih vokalnih načina izražavanja.
Hemijska komunikacija
[uredi | uredi izvor]Imaju odličan osećaj za mirise. Kada je uznemiren ili napadnut, u samoodbrani ispušta veoma neprijatan miris, nalik
onome kod tvora. Teritoriju obeležavaju urinom ili izmetom.
Vizuelna komunikacija
[uredi | uredi izvor]Kada su uznemireni i ljuti, uši im pocrvene i to postane vrlo uočljivo. Krv nadire u uši, a one su bez dlačica i imaju tanku kožu, što dodatno olakšava posmatraču da odredi u kakvom je raspoloženju ova životinja. Pošto su tasmanijski đavoli noćne životinje, tj. love noću, vid im je najjasniji u crno-beloj tehnici. Pod ovakvim uslovima obično zapažaju objekte koji se kreću, ali vrlo teško vide objekte koji miruju.
Način kretanja
[uredi | uredi izvor]Nije naročito sposoban za veranje po drveću, ali je dobar plivač i uspeva vešto da izbegne opasnost tako što u trenu sklizne ispod površine vode, a ukoliko ga gone, u stanju je da neprimetno pređe znatne razdaljine pre nego što izroni, obično na skrovitom mestu.[34]
Reprodukcija i briga o potomstvu
[uredi | uredi izvor]
Sezona parenja je u proleće i leto i najviše mladunaca se okoti u periodu od novembra do marta. Ženke dostižu polnu zrelost u drugoj godini života, fertilne su jednom godišnje.[35] Mužjaci se bore za njihovu naklonost, te se ženke pare sa dominantnim mužjakom. Parenje se odvija u martu, nezavisno od doba dana ili noći. Jednu ženku može pariti i više mužjaka, ako nije čuvana od strane dominantnog mužjaka. Kod ženki se oplodi više jajnih ćelija i na svet dolazi više mladunaca, ne samo jedan, kao kod većine torbara. Graviditet traje 21 dan i rađa se čak 20-ak sićušnih mladunaca koji teže samo od 0,18-0,24g.[36] Čim se rode, počinje kompeticija na putu do majčine torbe i bradavice. Ženka ima samo 4 bradavice, pa će tako samo 4 mladunca uspeti da prežive,[31] ostale će verovatno pojesti majka. U proseku preživi više malih ženki nego mužjaka.[35] Torba je okrenuta drugačije, ulaz je sa zadnje strane, te majka nema uvida u ono što se u torbi dešava sa mladuncima. Mladunci u torbi ostaju zakačeni za bradavicu i hrane se majčinim mlekom narednih 105 dana. Sisajući napreduju brzo, pa napuštaju torbe teže oko 500g. Nakon 15 dana uši su vidljive. Očni kapci se pojave nakon 16 dana, brkovi nakon 17 dana, usne nakon 20 dana. Mladima počne rasti krzno nakon oko 49 dana, a nakon 90 imaju celo krzno. Oči im se otvore kratko nakon razvitka krzna - između 87 i 93 dana.[35] U periodu od oktobra do decembra su još uvek u majčinoj blizini istražujući okolinu i žive sa njom u jazbini, a u januaru se potpuno osamostaljuju. Ženke odgajaju mlade šest mjeseci tokom jedne godine.
Reference
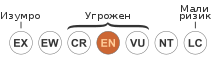
[uredi | uredi izvor]- ^ Hawkins, C.E.; McCallum, H.; Mooney, N.; Jones, M.; Holdsworth, M. (2008). „Sarcophilus harrisii”. IUCN Red List of Threatened Species. IUCN. 2008: e.T40540A10331066. doi:10.2305/IUCN.UK.2008.RLTS.T40540A10331066.en. Arhivirano iz originala 25. 03. 2019. g. Pristupljeno 27. 8. 2016. Listed as Endangered (EN A2be+3e v3.1)
- ^ Boitard, [Pierre] (n.d). „L'Ursin de Harris”. Le Jardin des plantes: Description et mœurs des mammifères de la Ménagerie et du Muséum d'histoire naturelle. Paris: Gustave Barba. str. 204.
- ^ Harris, G. P. (1807). „Description of two new Species of Didelphis from Van Diemen's Land”. Transactions of the Linnean Society of London. 9: 174—78. doi:10.1111/j.1096-3642.1818.tb00336.x.
- ^ Owen and Pemberton, p. 79.
- ^ a b v Owen and Pemberton, p. 8.
- ^ a b Groves, C.P. (2005). „Order Dasyuromorphia”. Ur.: Wilson, D.E.; Reeder, D.M. Mammal Species of the World: A Taxonomic and Geographic Reference (3rd izd.). Johns Hopkins University Press. str. 28. ISBN 978-0-8018-8221-0. OCLC 62265494.
- ^ Mitchell, Thomas (1839). Three Expeditions into the Interior, Volume II. T. & W. Boone. Online at Project Gutenberg Australia.
- ^ Stephenson, N. G. (1963). „Growth gradients among fossil monotremes and marsupials | The Palaeontological Association”. Palaeontology (na jeziku: engleski). 6 (4): 615—624.
- ^ Werdelin, L. (1987). „Some observations on Sarcophilus laniarius and the evolution of Sarcophilus”. Records of the Queen Victoria Museum, Launceston. 90: 1—27.
- ^ Owen and Pemberton, p. 7.
- ^ Krajewski, Carey; Driskell, Amy C.; Baverstock, Peter R.; Braun, Michael J. (1992). „Phylogenetic relationships of the thylacine (Mammalia:Thylacinidae) among dasyuroid marsupials: evidence from cytochrome b DNA sequences”. Proceedings of the Royal Society B: Biological Sciences. 250 (1327): 19—27. Bibcode:1992RSPSB.250...19K. PMID 1361058. S2CID 35414922. doi:10.1098/rspb.1992.0125.
- ^ a b v Owen and Pemberton, p. 34.
- ^ Krajewski, Carey; Westerman, Michael (2003). „Molecular Systematics of Dasyuromorpha”. Ur.: Jones, Menna; Dickman, Chris; Archer, Mike. Predators with Pouches: The Biology of Carnivorous Marsupials. Collingwood, Victoria: CSIRO Publishing. str. 16. ISBN 0-643-06634-9.
- ^ Long, John A.; Archer, Michael; Flannery, Timothy; Hand, Suzanne (2002). Prehistoric Mammals of Australia and New Guinea: One Hundred Million Years of Evolution. University of New South Wales Press. str. 55. ISBN 978-0-8018-7223-5.
- ^ Owen and Pemberton, p. 35.
- ^ a b v g d Owen and Pemberton, p. 36.
- ^ Owen and Pemberton, p. 37.
- ^ a b Owen and Pemberton, p. 38.
- ^ a b Owen and Pemberton, p. 39.
- ^ Owen and Pemberton, pp. 40–42.
- ^ „Completed genome is first step to tackling Tasmanian devil facial tumours” (Saopštenje). Wellcome Trust Sanger Institute. 16. 9. 2010. Pristupljeno 10. 12. 2015.
- ^ Tyndale-Biscoe, p. 143.
- ^ a b v Jones, M. E.; Paetkau, David; Geffen, Eli; Moritz, Craig (2004). „Genetic diversity and population structure of Tasmanian devils, the largest marsupial carnivore”. Molecular Ecology. 13 (8): 2197—2209. Bibcode:2004MolEc..13.2197J. PMID 15245394. S2CID 14068282. doi:10.1111/j.1365-294X.2004.02239.x.
- ^ Morris, K.; Austin, J. J.; Belov, K. (5. 12. 2012). „Low major histocompatibility complex diversity in the Tasmanian devil predates European settlement and may explain susceptibility to disease epidemics”. Biology Letters. 9 (1): 20120900. PMC 3565505
 . PMID 23221872. doi:10.1098/rsbl.2012.0900.
. PMID 23221872. doi:10.1098/rsbl.2012.0900.
- ^ Lachish, S.; Miller, K.; Storfer, A.; Goldizen, A.; et al. (2010). „Evidence that disease-induced population decline changes genetic structure and alters dispersal patterns in the Tasmanian devil”. Heredity. 106 (1): 172—182. PMC 3183847 . PMID 20216571. doi:10.1038/hdy.2010.17.
- ^ „Sarcophilus harrisii – Tasmanian Devil”. Species Profile and Threats Database. Department of Sustainability, Environment, Water, Population and Communities. Pristupljeno 30. 9. 2010.
- ^ „Draft Recovery Plan for the Tasmanian devil (Sarcophilus harrisii)” (PDF). Hobart: Department of Primary Industries, Parks, Water and Environment. 2010. Pristupljeno 3. 9. 2015.
- ^ a b v Siddle, Hannah V.; Marzec, Jolanta; Cheng, Yuanyuan; Jones, Menna; et al. (2010). „MHC gene copy number variation in Tasmanian devils: Implications for the spread of a contagious cancer”. Proceedings of the Royal Society B. 277 (1690): 2001—06. PMC 2880097 . PMID 20219742. doi:10.1098/rspb.2009.2362.
- ^ Epstein, Brendan; Jones, Menna; Hamede, Rodrigo; Hendricks, Sarah; McCallum, Hamish; Murchison, Elizabeth P.; Schönfeld, Barbara; Wiench, Cody; Hohenlohe, Paul; Storfer, Andrew (30. 8. 2016). „Rapid evolutionary response to a transmissible cancer in Tasmanian devils”. Nature Communications. 7: 12684. Bibcode:2016NatCo...712684E. PMC 5013612 . PMID 27575253. doi:10.1038/ncomms12684.
- ^ Guiler, E.R. 1983. Tasmanian Devil in R. Strahan Ed. The Australian Museum Complete Book of Australian Mammals. p 27–28. Angus & Robertson. ISBN 978-0-207-14454-7.
- ^ a b Department of Primary Industries, Water and Environment. Tasmanian Devil - Frequently Asked Questions
- ^ Wroe, S, McHenry, C, and Thomason, J. 2005. Proceedings of the Royal Society B-Biological Sciences 272:619–625 „Bite club: comparative bite force in big biting mammals and the prediction of predatory behaviour in fossil taxa”. PMID 15817436.
- ^ Britannica, Encyclopædia. „Tasmanian devil (marsupial)”. Britannica.com. Pristupljeno 16. 3. 2010.
- ^ Sanderson 1967.
- ^ a b v Guiler, E.R. 1970. Observations on the Tasmanian devil, Sarcophilus harrisii II. Reproduction, Breeding and Growth of Pouch Young. Australian Journal of Zoology 18:63–70
- ^ Fisher, D.O. et al. 2001. The ecological basis of life history variation in marsupials, Appendix A. Ecology 82:3531–3540
Literatura
[uredi | uredi izvor]- Boitard, [Pierre] (n.d). „L'Ursin de Harris”. Le Jardin des plantes: Description et mœurs des mammifères de la Ménagerie et du Muséum d'histoire naturelle. Paris: Gustave Barba. str. 204.
- Guiler, ER (1992). The Tasmanian devil. Hobart, Tasmania: St David's Park Publishing. ISBN 978-0-7246-2257-3.
- Figueroa, Don; Furman, Simon; Yee, Ben; Khanna, Dan M.; Guidi, Guido; Isenberg, Jake; Matere, Marcelo; Roche, Roche; Ruffolo, Rob; Williams, Simon (2008). The Transformers Beast Wars Sourcebook. San Diego, California: IDW Publishing. ISBN 978-1-60010-159-5.
- Owen, D; Pemberton, David (2005). Tasmanian Devil: A unique and threatened animal. Crows Nest, New South Wales: Allen & Unwin. ISBN 978-1-74114-368-3. Pristupljeno 22. 8. 2010.
- Paddle, Robert (2000). The Last Tasmanian Tiger: The History and Extinction of the Thylacine. Oakleigh, Victoria: Cambridge University Press. ISBN 978-0-521-53154-2.
- Tyndale-Biscoe, Hugh (2005). Life of marsupials. Collingwood, Victoria: CSIRO Publishing. ISBN 978-0-643-06257-3.
- Hesterman, H.; Jones, S. M.; Schwarzenberger, F. (2008). „Pouch appearance is a reliable indicator of the reproductive status in the Tasmanian devil and the spotted-tailed quoll”. Journal of Zoology. 275 (2): 130—138. doi:10.1111/j.1469-7998.2008.00419.x.
- McDonald-Madden, E.; Probert, W. J. M.; Hauser, C. E.; Runge, M. C.; Possingham, H. P.; Jones, M. E.; Moore, J. L.; Rout, T. M.; Vesk, P. A.; Wintle, B. A. (2010). „Active adaptive conservation of threatened species in the face of uncertainty” (PDF). Ecological Applications. 20 (5): 1476—1489. PMID 20666263. doi:10.1890/09-0647.1.
Spoljašnje veze
[uredi | uredi izvor]- Parks and Wildlife Tasmania – Tasmanian Devil – vocalisation, movie, FAQ
- Save the Tasmanian Devil Архивирано на сајту Wayback Machine (21. март 2009) – Tasmanian government conservation program
- View the Tasmanian devil genome in Ensembl
| Државне | |
|---|---|
| Остале | |