
Липид


| Типови масти у храни |
|---|
|
| Види још |
Липиди (грч. λίπος — маст) јесу једињења различитог састава по правилу нерастворна у води, а растворна у органским растварачима.[4] Биолошки су веома значајна једињења. Они су основна компонента биолошких мембрана и утичу на њихову пропустљивост, учествују у предаји нервних импулса, стварају контакте међу ћелијама, чине енергетске резерве, штите организам од механичких повреда и формирају термоизолациони слој.[5]
Због специфичне хемијске структуре, липиди се састоје од неполарног и поларног дела. Они су често упрошћено називани мастима, али тај назив није исправан јер су липиди група која не обухвата само једињења масних киселина и глицерола, већ и других супстанци. Дакле, масти су у ствари само једна од неколико група липида.[6]
Преглед
[уреди | уреди извор]Липиди обухватају велику групу разнородних једињења са заједничком одликом нерастворљивости у води, и растворљивости у органским растварачима: бензолу, хлороформу, ацетону, врелом алкохолу. Липиди у биљним и животињским организмима учествтвују као градивне и резервне материје. У метаболизму липида значајну улогу има активирана сирћетна киселина (или ацетил-коензим-ацетил-КоА). Према структурној организацији, масти се деле на:
- просте и
- сложене масти.[7]
Прости липиди су естри трихидроксилног алкохола (глицерола) и масних киселина (олеинске, памитинске и стеаринске). Чврсте масти (лој) чине једињења глицерола са засићеном палмитинском (C16H36O2) или стеаринском (C18H36O2) киселином. За разлику од њих, у састав течних масти или уља улази олеинска (C17H34O2). Просте масти са водом чине нестабилне емулзије.
Загревањем масти у присуству алкалија (NaOH или KOH) долази до сапонификације, чиме се добијају једињења сапуна (која су растворна у води). Процесом сапонификације масти се разграђују на глицерол и соли масних киселина. Увођењем водоника у незасићене масне киселине (процесом хидрогенизације), оне прелазе у засићене са измењеним мирисом и укусом. Захваљујући примени овог поступка, у индустрији се развио поступак прављења маргарина и биљног масла.
Липиди, као резервна органску материја, су карактеристични по великој енергетској вредност, а почињу се трошити само у недостатку глукозе. Вишак угљених хидрата у организму се претвара у масти и складишти у различитим органима (код животиња, нпр. најчешће се одлажу поткожно). Такав случај је посебно значајан код птица селица и животиња са сезонским зимским сном.
Воскови, за разлику од глицетида, уместо глицерола садрже дуголанчане једновалентне алкохоле. Поред алкохола, садрже и више масне киселине. Заступљени су у биљкама и животињама. Нису растворљиви у води, већ у органским растварачима. Међу познатијим супстанцама ове групе су: пчелињи восак, спермацет и ланолин.
У сложене липиде убрајају се фосфолипиди, гликолипиди и липопротеини. Изграђени су од глицерола, масних киселина и фосфорне киселине.[7]
Од познатијих фосфолипида, најчешћи су лецитин, кефалин, плазмалоген и лизофосфолипид. Фосфолипиди чине темељну компоненту живе ћелијске мембране и осталих ћелијских улдопина. Одвијање нормалних животних процеса у организму, везано је за специфичне функције сложених липида. Тако је за контраховање глатке мускулатуре неопходан медијатор ацетилхолин. Он такође има посебан значај у подраживању парасимпатичких нерава, те снижавање крвног притиска. Фосфолипиди такође учествују у пермеабилности ћелијских (живих) мембрана, посебно при преносу јона Na+ и K+ у срчаном мишићу (миокарду), док од неких других зависи пермеабилност мембране митохондрија. Лизофосфатиди изазивају хемолизу (разградњу) еритроцита. У змијском отрову налази се фосфолипаза А, због чега након уједа долази до хемолизе еритроцита.
Гликолипиди обухватају цереброзиде, ганглиозиде, сулфатиде итд. Цереброзиди су први пут изоловани из мозга (cerebrum), по чему су и добили име. Ганглиозиди су изоловани из ганглијских ћелија мозга (сиве масе). Сулфатиди су једињења сложених липида изолована из јетре и бубрега, а релативно високим садржајем ових гликолипида обогаћен је и мозак. Сва ова једињења у организмима животиња имају врло специфичне и значајне функције.[7]
Класификација и особине
[уреди | уреди извор]Прости липиди су супстанце чији се молекули састоје само од остатака масних киселина и алкохола (најчешће глицерола). Овде спадају масти и уља (триглицериди) и воскови.[8][9]
Сложени липиди укључују деривате фосфорне киселине (фосфолипиди) и липиде који садрже остатке угљених хидрата (гликолипиди). Овде спадају и стероиди.[10]
Ако се животињска или биљна ткива третирају органским растварачима, део ће се растворити. Компоненте раствора се називају липиди. Липидна фракција садржи супстанце различитих типова.
Масне киселине спадају у алифатичне карбоксилне киселине. Из липида је изоловано више од 70 разних масних киселина. Деле се на засићене и незасићене. Природне масне киселине садрже паран број C атома, а низ није разгранат, већ је нормалан. Испод су набројане неке од најважнијих масних киселина (масне се зову зато што се налазе у мастима):
| Засићене масне киселине | |
| Тривијални назив | Структурна формула |
| бутерна | CH3(CH2)2COOH |
| капронска | CH3(CH2)4COOH |
| каприлна | CH3(CH2)6COOH |
| капринска | CH3(CH2)8COOH |
| лауринска | CH3(CH2)10COOH |
| миристинска | CH3(CH2)12COOH |
| палмитинска | CH3(CH2)14COOH |
| стеаринска | CH3(CH2)16COOH |
| арахидинска | CH3(CH2)18COOH |
| бегенска | CH3(CH2)20COOH |
| лигноцеринска | CH3(CH2)22COOH |
| Незасићене масне киселине | |
| Тривијални назив | Структурна формула |
| кротонска | CH3 - CH = CH - COOH |
| палматоолеинска | CH3 - (CH2)5 - CH = CH - (CH2)7 - COOH |
| олеинска | CH3 - (CH2)7 - CH = CH - (CH2)7 - COOH |
| елаидинска | CH3 - (CH2)7 - CH = CH - (CH2)11 - COOH |
| нервонска | CH3 - (CH2)7 - CH = CH - (CH2)13 - COOH |
| линолна | CH3 - (CH2)4 - CH = CH - CH2 - CH = CH - (CH2)7 - COOH |
| линолеинска | CH3 - CH2 - CH = CH - CH2 - CH = CH - CH2 - CH = CH - (CH2)7 - COOH |
| арахидонска | CH3 - (CH2)4 - CH = CH - CH2 - CH = CH - CH2 - CH = CH - CH2 - CH = CH - (CH2)3 - COOH |
| клупадонска | CH3 - CH2 - CH = CH - (CH2)2 - CH = CH - CH2 - CH = CH - (CH2)2 - CH = CH - (CH2)2 - CH = CH - (CH2)2 - COOH |
Најважније природне масне киселине садрже 16 или 18 C атома. Незасићене масне киселине обично садрже двоструке везе између C9 и C10. Остале двоструке везе нису конјуговане и налазе се према CH3- крају низа. Имају цис- конфигурацију, а веома ретко транс конфигурацију. Масне киселине са већим бројем C атома су практично нерастворне у води. Њихове Na и K- соли се зову сапуни. Оне образују мицеле у води код којих хе негативан COO- крај окренут ка води, а неполаран C- низ је окренут ка унутрашњости мицеле. У целини мицеле су наелектрисане негативно и у раствору остају суспендоване јер се међусобно одбијају.
Глицериди
[уреди | уреди извор]Неутралне масти (глицериди) (ацилглицериди) су естри глицерола (глицерина) и масних киселина. У односу на број естарских веза у молекулу разликују се моноглицериди, диглицериди и триглицериди (односно, моноацилглицерини, диацилглицерини и триацилглицерини).
Масти се у организмима налазе као протоплазматичне масти (у одређеној количини) и као резервне масти (у неодређеној количини). Основну масу масти чине триглицериди. Ако су сва три киселинска остатка иста, прости триглицериди (на пример, трипалмитин), а ако су остаци од од различитих киселина, мешовити триглицериди. Код давања назива, цифре указују на коме се C атому налази одговарајући киселински остатак (на пример. 1-олео-2-пал-митостеарин).
Уколико су киселински остаци дужи, утолико расте и тачка топљења. Ако су киселине незасићене, тачка топљења са бројем двоструких веза опада. Животињске масти садрже знатну количину засићених масних киселина (нарочито стеаринску и палмитинску) па су на собној температури чврсте. Ако се у молекулу налази доста незасићених киселина то су онда уља, која су на собној температури течна (маслиново уље, сојино, сунцокретово, уље уљане репице). Маст код људи има тачку топљења 15 °C и садржи 70% олеинске киселине.
Глицериди реагују као естри, а посебно је важна реакција хидролизе, при чему се издваја глицерин и масне киселине. Хидролиза се може вршити ензимски или дејством киселина или база. Базна хидролиза са натријум хидроксидом (NaOH) или са калијум хидроксидом (KOH) се користи за производњу сапуна (реакција сапонификације, сапо-сапун).
Масти садрже разне примесе које се уклањају, а заостаје тзв. неутрална маст. Она се карактерише:
- јодним бројем (број грама јода кога везује 100 g масти под одређеним условима и који карактерише степен незасићености. Јодни број свињске масти је 46-66, а маслиновог уља око 85).
- киселински број (број mg KOH потребан за неутрализацију 1 g масти и који указује на количину слободних масних киселина у масти).
- сапонификациони број (број mg KOH потребан за неутрализацију свих киселина – и слободних и везаних у триглицериде – у 1 g масти).
Воскови
[уреди | уреди извор]Воскови су естри виших масних киселина и виших алкохола са 20 до 70 C атома у молекулу. Они се налазе у саставу масти која покрива кожу, вуну, перје. Код биљака су 80% свих липида воскови који стварају заштитни слој на површини биљке и штите је од испаравања воде.
Природни восак (пчелињи, ланолин) поред поменутих естара садржи и малу количину слободних масних киселина, неких алкохола и угљоводоника, са 20-35 C атома у молекулу. Општа формула воскова је:
Фосфолипиди
[уреди | уреди извор]Фосфолипиди или фосфатиди се деле на глицерофосфолипиде и сфинголипиде.
- Глицерофосфолипиде су деривати фосфатидне киселине и садрже глицерол, масне киселине, фосфатидну киселину и азотна једињења. Њихова општа формула је:

где су R и R' остаци виших масних киселина, а R" је остатак неког азотног једињења и фосфорне киселине.
Један део молекула (R и R') је јако хидрофобан, а други део је хидрофилан (негативно наелектрисање у фосфорној киселини и позитивно наелектрисање азотног једињења – R"). Од свих липида глицерофосфолипиди су најполарнији, а у воденој средини се налази у облику мицела. Има неколико група глицерофосфолипида:
- Фосфатидилхолини, лецитини садрже глицерол, више масне киселине, фосфорну киселину и холин:

- Фосфатидилетаноламини, кефалини уместо холина садрже етаноламин:

Горње две групе су главна липидна компонента мембране ћелија.
- Фосфатидилсерини као азотна једињења садрже остатак серина:

Од масних киселина које улазе у састав глицерофосфолипида најчешће су стеаринска, палмитинска, олеинска и линоленска. При њиховој хидролизи (отров кобре) издваја се остатак незасићене масне киселине, а преостало једињење има јако хемолитичко дејство.
Сфинголипиди
[уреди | уреди извор]- Сфинголипиди не садрже глицерол већ алкохол сфингозин:
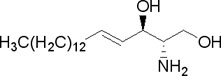

Најчешће су сфингомијелини који се налазе у мембранама ћелија и то посебно нервних. Хидролизом дају један молекул масне киселине, један молекул незасићеног двовалентног алкохола сфингозина, један молекул азотног једињења (најчешће холина) и један молекул фосфорне киселине. Хемијска формула сфингомијелина дата је испод:

Поларни део је наелектрисан и позитивно (холин) и негативно (остатак фосфорне киселине), а остали део молекула је неполаран.
Гликолипиди
[уреди | уреди извор]Гликолипиди су сложени липиди који садрже остатак угљених хидрата (најчешће D-галактозе). Битни су за рад биолошких мембрана. Налазе се у мозгу и крви. Деле се не две групе:
- Цереброзиди садрже хексозу (често D-галактозу) везану етарском везом са сфингозином, а садрже и неку масну киселину са 24 C атома (лигноцеринска, церебронска):

Много цереброзида се налази у мембранама нервних ћелија.
Ганглиозиди
[уреди | уреди извор]Ганглиозиди хидролизом дају вишу масну киселину, сфингозин, D-глукозу, D-галактозу као и деривате аминошећера: N-ацетилглукозамин и N-ацетилнеураминску киселину. Налазе се претежно у сивој кори мозга и мембранама нервих ћелија.
Сви горе поменути липиди спадају у сапонификујуће липиде јер при њиховој базној хидролизи настају сапуни. Постоје и липиди који се не хидролизују уз ослобађање масних киселина. У ту групу спадају стероиди.
Биолошке функције
[уреди | уреди извор]Мембране
[уреди | уреди извор]
Еукариотске ћелије су компартментализоване у органеле обавијене мембранама, које врше различите биолошке функције. Глицерофосфолипиди су главна структурна компонента биолошких мембрана, као што су ћелијска мембрана и интраћелијске мембране органела; у животињским ћелијама мембране физички одвајају интраћелијске компоненте од екстрацелуларног окружења. Глицерофосфолипиди су амфипатички молекули (садрже хидрофобне и хидрофилне регионе) који имају глицеролно језгро везано са две масне киселине естарским везама, и за једну „чеону“ групу путем фосфатне естарске везе. Док су глицерофосфолипиди главне компоненте биолошких мембрана, друге неглицеридне липидне компоненте као што су сфингомијелин и стероли (углавном холестерол у животињским ћелијским мембранама) су исто тако присутни у биолошким мембранама.[11] Код биљки и алги галактозилдиацилглицероли[12] и сулфохиновозилдиацилглицерол,[13] којима недостаје фосфатна група, су важне компоненте мембрана хлоропласта и сродних органела и најзаступљенији су липиди у фотосинтетичким ткивима, укључујући она код виших биљки, алги и појединих бактерија.
Највећа липидна компонента биљне тилакоидне мембране је моногалактозил диглицерид (МГДГ) који обично не формира двослој, док су фосфолипиди присутни у мањој мери. Упркос тој јединственој липидној композицији, за хлоропластне тилакоидне мембране је путем магнетне резонанце и електронске микроскопије показано да садрже динамичку липидни двослојну матрицу.[14]
Биолошка мембрана је форма ламеларне фазе липидног двослоја. Формирање липидног двослоја је енергетски преферентан процес кад су горе описани глицерофосфолипиди у воденом окружењу.[15] То је познато као хидрофобни ефекат. У воденој средини, поларне главе липида се поравнавају ка поларном, воденом окружењу, док хидрофобни репови минимизују свој контакт са водом и групишу се заједно, формирајући везикуле; у зависности од концентрације липида, та биофизичка интеракција може да доведе до формирања мицела, липозома, или липидних двослоја. Други облици агрегације су исто тако примећени и формирају део полиморфизма амфифилног (липидног) понашања. Фазно понашање је област изучавања у оквиру биофизике и предмет је текућих академских истраживања.[16][17] Мицеле и двослојеви се формирају у поларном медијуму процесу познатом као хидрофобни ефекат.[18] При растварању липофилне или амфифилне супстанце у поларној средини, поларни молекули (и.е., вода у воденим растворима) постају у већој мери уређени око растворене липофилне супстанце, пошто поларни молекули не могу да формирају водоничне везе са липофилним областима амфифила. Стога у воденом окружењу, молекули воде формирају уређене "клатратне" кавезе око растворених липофилних молекула.[19]
Организовање липида у протоћелијске мембране је кључни корак у моделима абиогенезе, порекла живота.[20]
Чување енергије
[уреди | уреди извор]Триглицериди, ускладиштени у адипозном ткиву, су главна форма енергетског складишта код животиња и биљки. Адипоцит, или масна ћелија, је дизајниран за континуирану синтезу и разлагање триглицерида код животиња, при чему се разлагање углавном активира ензимима сензитивним на хормоне, липазама.[21] Комплетна оксидација масних киселина пружа висок калоријски садржај, око 9 kcal/g, у поређењу са 4 kcal/g за разлагање угљених хидрата и протеина. Миграторне птице могу да лете преко великих растојања без конзумирања хране користећи енергију сачувану у облику триглицерида као гориво за свој лет.[22]
Сигнализација
[уреди | уреди извор]Задњих година је постало јасно да је липидна сигнализација витални део ћелијске сигнализације.[23][24] Липидна сигнализација се може одвијати путем активације Г протеин спрегнутих или нуклеарних рецептора, и чланови неколико различитих категорија липида су идентификовани као сигнални молекули и ћелијски гласници.[25] Међу њима су сфингозин-1-фосфат, сфинголипид изведен из керамида који је потентан гласнички молекул који учествује у регулацији калцијумске мобилизације,[26] у ћелијском расту, и апоптози;[27] диацилглицерол (ДАГ) и фосфатидилинозитол фосфати (ПИП), који учествују у калцијумом посредованој активацији протеинске киназе Ц;[28] простагландини, који су један од типова из масних киселина изведених еикосаноида који учествују у инфламацији и имуности;[29] стероидни хормони као што су естроген, тестостерон и кортизол, који модулишу домаћинове функције репродукције, метаболизма и крвног притиска; и оксистероли као што је 25-хидрокси-холестерол који је агонист јетреног X рецептора.[30] Познато је да фосфатидилсерински липиди учествују у сигнализацији фагоцитозе апоптотичких ћелија и/или делова ћелија. Они остварују тај циљ тако што су изложени на екстрацелуларном лицу ћелијске мембране након инактивације флипаза које их ексклузивно постављају на цитозолну страну и активације скрамблаза, које изокрећу оријентацију фосфолипида. Након тога друге ћелије препознају фосфатидилсерине и фагоцитозирају ћелије или ћелијске фрагменте који их излажу.[31]
Друге функције
[уреди | уреди извор]Витамини растворни у мастима (А, Д, Е и К) – који су на изопрену базирана једињења – су есенцијални нутријенти који се чувају у јетри и масним ткивима, са разноврсним опсегом функција. Ацил-карнитини учествују у транспорту и метаболизму масних киселина „у и из“ митохондрија, где они подлежу бета оксидацији.[32] Полипреноли и њихови фосфорилисани деривати такође имају важну транспортне улоге, у том случају у транспорту олигосахарида кроз мембране. Полипренолни фосфатни шећери и полипренолни дифосфатни шећери узимају учешћа у реакцијама гликозилације изван цитоплазме, у биосинтези екстрацелуларних полисахарида (на пример, пептидогликанској полимеризацији код бактерија), и у N-гликозилацији еукариотских протеина.[33][34] Кардиолипини су поткласа глицерофосфолипида који садрже четири ацилна ланца и три глицеролне групе, која је посебно изобилна у унутрашњој митохондријској мембрани.[35][36] Сматра се да они активирају ензиме који учествују у оксидативној фосфорилацији.[37] Липиди исто тако формирају основу стероидних хормона.[38]
Метаболизам
[уреди | уреди извор]Главни прехрамбени липиди код људи и животиња су биљни триглицериди, стероли, и мембрански фосфолипиди. Процесом липидног метаболизма се синтетишу и деградирају ускладиштени липиди и производе се структурни и функционални липиди карактеристични за појединачна ткива.
Биосинтеза
[уреди | уреди извор]Код животиња, кад постоји вишак понуде прехрамбених угљених хидрата, сувишни угљени хидрати се конвертују у триглицериде. Тај процес обухвата синтезу масних киселина из ацетил-КоА и естерификацију масних киселина ради продукције триглицерида. Овај процес се назива липогенеза.[39] Масне киселине се формирају синтезом масних киселина, којом се полимеризују и затим редукују ацетил-КоА јединице. Ацилни ланци масних киселина се продужавају циклусом реакција које додају ацетил групу, редукују је до алкохола, дехидратишу алкохол до алкенске групе и затим се поново врши редукција до алканске групе. Ензими масно киселинске биосинтезе се деле у две групе, код животиња и гљива све реакције синтезе масних киселина се изводе посредством једног мултифункционалног протеина,[40] док у биљним пластидима и бактеријама засебни ензими врше сваки корак тог процеса.[41][42] Масне киселине се могу накнадно конвертовати до триглицерида који се пакују у липопротеинима и излучују из јетре.
Синнтеза незасићених масних киселине обухвата реакцију десатурације, при чему се двострука веза уводи у масни ацилни ланац. На пример, код људи, десатурација стеаринске киселине посредством ензима стеароил-КоА десатураза-1 производи олеинску киселину. Двоструко незасићена масна киселина линолеинска киселина, као и троструко незасићена α-линолеинска киселина се не могу синтетисати у ткивима сисара, и стога су есенцијалне масне киселине и морају се уносити као храна.[43]
Триглицеридна синтеза се одвија у ендоплазматичном ретикулуму путем метаболичких путева у којима се ацилне групе у масним ацил-КоА молекулима преносе на хидроксине групе глицерол-3-фосфата и диацилглицерола.[44]
Терпени и изопреноиди, укључујући каротеноиде, се формирају путем прављења и модификације изопренских јединица из реактивних прекурзора изопентенил пирофосфата и диметилалил пирофосфата.[45] Ти прекурзори могу да буду направљени на различите начине. Код животиња и археја, мевалонатни путем се формирају та једињења из ацетил-КоА,[46] док се у биљкама и бактеријама у немевалонатном путу користе пyруват и глицералдехид 3-фосфат као супстрати.[45][47] Једна важна реакција која користи те активиране изопренске доноре је стероидна биосинтеза. Овде се изопренске јединице спајају да би се формирао сквален и затим се савијају да би настала група прстенова који сачињавају ланостерол.[48] Ланостерол се може даље конвертовати у друге стероиде као што је холестерол и ергостерол.[48][49]
Деградација
[уреди | уреди извор]Бета оксидација је метаболички процес којим се масне киселине разлажу у митохондријама и/или у пероксизомима да би се формирао ацетил-КоА. Највећим делом, масне киселине се оксидују путем механизма који је сличан са, мада не и идентичан са, реверзним процесом синтезе масних киселина. Другим речима, двоугљенични фрагменти се уклањају секвенцијално са карбоксилног краја киселине након корака дехидрогенације, хидрације, и оксидације уз формирање бета-кето киселине, које се одваја тиолизом. Ацетил-КоА се затим ултиматно конвертује у АТП, CO2, и Х2О користећи циклус лимунске киселине и ланац електронског транспорта. Циклус лимунске киселине може да почне од ацетил-КоА кад се масноћа разлаже ради енергије, ако је присутно мало или је одсутна глукоза. Енергетски принос од комплетне оксидације масне киселине палмитата је 106 АТП.[50] За деградацију незасићених и непарних масних киселина неопходни су додатни ензиматски процеси.
Референце
[уреди | уреди извор]- ^ Maitland 1998, стр. 139
- ^ Stryer et al.. стр. 328.
- ^ Stryer et al.. стр. 330.
- ^ Clayden, Jonathan; Greeves, Nick; Warren, Stuart; Wothers, Peter (2001). Organic Chemistry (I изд.). Oxford University Press. ISBN 978-0-19-850346-0.
- ^ David L. Nelson; Michael M. Cox (2005). Principles of Biochemistry (IV изд.). New York: W. H. Freeman. ISBN 0-7167-4339-6.
- ^ Шербан 2010.
- ^ а б в Međedović, S.; Maslić, E.; Hadžiselimović, R. (2002). Biologija 2. Sarajevo: Svjetlost. ISBN 978-9958-10-222-6.
- ^ Кораћевић, Даринка; Бјелаковић, Гордана; Ђорђевић, Видосава. Биохемија. Савремена администрација. ISBN 978-86-387-0622-8.
- ^ аутора, Група (2006). Угрешић, Ненад, ур. Фармакотерапијски водич 3 (PDF). Београд. ISSN 1451-4680. Архивирано из оригинала (PDF) 15. 07. 2011. г. Приступљено 23. 05. 2010.
- ^ Keith Parker; Laurence Brunton; Goodman, Louis Sanford; Lazo, John S.; Gilman, Alfred (2006). Goodman & Gilman's The Pharmacological Basis of Therapeutics (11. изд.). New York: McGraw-Hill. ISBN 0071422803.
- ^ Stryer et al.. стр. 329–331.
- ^ Heinz E. "Plant glycolipids: structure, isolation and analysis". стр. 211–332 in Advances in Lipid Methodology, Vol. 3. W.W. Christie (ed.). Oily Press. Dundee. 1996. ISBN 978-0-9514171-6-4.
- ^ Hölzl, G.; Dörmann, P. (2007). „Structure and function of glycoglycerolipids in plants and bacteria”. Progress in Lipid Research. 46 (5): 225—243. PMID 17599463. doi:10.1016/j.plipres.2007.05.001.
- ^ Yashroy, RC. (1990). „Magnetic resonance studies of dynamic organisation of lipids in chloroplast membranes”. Journal of Biosciences. 15 (4): 281—288. doi:10.1007/BF02702669.
- ^ Stryer et al.. стр. 333–334.
- ^ van Meer G, Voelker DR, Feigenson GW (2008). „Membrane lipids: where they are and how they behave”. Nature Reviews Molecular Cell Biology. 9 (2): 112—124. PMC 2642958
 . PMID 18216768. doi:10.1038/nrm2330.
. PMID 18216768. doi:10.1038/nrm2330.
- ^ Feigenson, GW. (2006). „Phase behavior of lipid mixtures”. Nature Chemical Biology. 2 (11): 560—563. PMC 2685072 . PMID 17051225. doi:10.1038/nchembio1106-560.
- ^ Wiggins, PM. (1990). „Role of water in some biological processes”. Microbiological Reviews. 54 (4): 432—449. PMC 372788 . PMID 2087221.
- ^ Raschke, T. M.; Levitt, M. (2005). „Nonpolar solutes enhance water structure within hydration shells while reducing interactions between them”. Proceedings of the National Academy of Sciences of the United States of America. 102 (19): 6777—6782. PMC 1100774 . PMID 15867152. doi:10.1073/pnas.0500225102.
- ^ Segré, D.; Ben-Eli D; Deamer, D.; Lancet D (2001). „The Lipid World” (PDF). Origins of Life and Evolution of Biospheres. 31 (1–2): 119—145. PMID 11296516. doi:10.1023/A:1006746807104. Архивирано из оригинала (PDF) 11. 09. 2008. г. Приступљено 13. 02. 2017.
- ^ Brasaemle, DL. (2007). „Thematic review series: adipocyte biology. The perilipin family of structural lipid droplet proteins: stabilization of lipid droplets and control of lipolysis”. Journal of Lipid Research. 48 (12): 2547—2559. PMID 17878492. doi:10.1194/jlr.R700014-JLR200.
- ^ Stryer et al.. стр. 619.
- ^ „Lipid”. Current Opinion in Plant Biology. 7 (3): 329—236. 2004. PMID 15134755. doi:10.1016/j.pbi.2004.03.012.
|first1=захтева|last1=у Authors list (помоћ) - ^ Dinasarapu, A. R.; Saunders, B.; Ozerlat I; Azam K, Subramaniam S (2011). „Signaling gateway molecule pages—a data model perspective”. Bioinformatics. 27 (12): 1736—1738. PMC 3106186 . PMID 21505029. doi:10.1093/bioinformatics/btr190.
- ^ Eyster, KM. (2007). „The membrane and lipids as integral participants in signal transduction”. Advances in Physiology Education. 31 (1): 5—16. PMID 17327576. doi:10.1152/advan.00088.2006.
- ^ Hinkovska-Galcheva V; VanWay, S. M.; Shanley, T. P.; Kunkel, R. G. (2008). „The role of sphingosine-1-phosphate and ceramide-1-phosphate in calcium homeostasis”. Current Opinion in Investigational Drugs. 9 (11): 1191—1205. PMID 18951299.
- ^ Saddoughi, S. A.; Song, P.; Ogretmen, B. (2008). „Roles of bioactive sphingolipids in cancer biology and therapeutics”. Subcellular Biochemistry. Subcellular Biochemistry. 49: 413—440. ISBN 978-1-4020-8830-8. PMC 2636716 . PMID 18751921. doi:10.1007/978-1-4020-8831-5_16.
- ^ Klein, C.; Malviya, A. N. (2008). „Mechanism of nuclear calcium signaling by inositol 1,4,5-trisphosphate produced in the nucleus, nuclear located protein kinase C and cyclic AMP-dependent protein kinase”. Frontiers in Bioscience. 13 (13): 1206—1226. PMID 17981624. doi:10.2741/2756.
- ^ Boyce, JA. (2008). „Eicosanoids in asthma, allergic inflammation, and host defense”. Current Molecular Medicine. 8 (5): 335—349. PMID 18691060. doi:10.2174/156652408785160989.
- ^ „Liver”. Cardiovascular Therapy. 26 (4): 297—316. 2008. PMID 19035881. doi:10.1111/j.1755-5922.2008.00062.x.
|first1=захтева|last1=у Authors list (помоћ) - ^ Biermann, M.; Maueröder C; Brauner JM,Chaurio R, Janko C, Herrmann M, Muñoz LE (2013). „Surface code—biophysical signals for apoptotic cell clearance”. Physical Biology. 10 (6): 065007. PMID 24305041. doi:10.1088/1478-3975/10/6/065007.
- ^ Indiveri, C; Tonazzi, A; Palmieri, F (1991). „Characterization of the unidirectional transport of carnitine catalyzed by the reconstituted carnitine carrier from rat liver mitochondria”. Biochimica et biophysica acta. 1069 (1): 110—6. PMID 1932043.
- ^ Parodi, A. J.; Leloir, L. F. (1979). „The role of lipid intermediates in the glycosylation of proteins in the eucaryotic cell”. Biochimica et Biophysica Acta. 559 (1): 1—37. PMID 375981. doi:10.1016/0304-4157(79)90006-6.
- ^ Helenius, A.; Aebi M (2001). „Intracellular functions of N-linked glycans”. Science. 291 (5512): 2364—2369. PMID 11269317. doi:10.1126/science.291.5512.2364.
- ^ Nowicki, M.; Frentzen M (2005). „Cardiolipin synthase of Arabidopsis thaliana”. FEBS Letters. 579 (10): 2161—2165. PMID 15811335. doi:10.1016/j.febslet.2005.03.007.
- ^ Gohil, V. M.; Greenberg, M. L. (2009). „Mitochondrial membrane biogenesis: phospholipids and proteins go hand in hand”. Journal of Cell Biology. 184 (4): 469—472. PMC 2654137 . PMID 19237595. doi:10.1083/jcb.200901127.
- ^ Hoch, FL. (1992). „Cardiolipins and biomembrane function”. Biochimica et Biophysica Acta. 1113 (1): 71—133. PMID 10206472. doi:10.1016/0304-4157(92)90035-9.
- ^ Steroids Архивирано на сајту Wayback Machine (23. октобар 2011). Elmhurst.edu. Приступљено 2013-10-10.
- ^ Stryer et al.. стр. 634.
- ^ Chirala, S.; Wakil S (2004). „Structure and function of animal fatty acid synthase”. Lipids. 39 (11): 1045—1053. PMID 15726818. doi:10.1007/s11745-004-1329-9.
- ^ White, S. W.; Zheng, J.; Zhang, Y. M.; Rock, C. O. (2005). „The structural biology of type II fatty acid biosynthesis”. Annual Review of Biochemistry. 74: 791—831. PMID 15952903. doi:10.1146/annurev.biochem.74.082803.133524.
- ^ Ohlrogge, J.; Jaworski J (1997). „Regulation of fatty acid synthesis”. Annual Review of Plant Physiology and Plant Molecular Biology. 48: 109—136. PMID 15012259. doi:10.1146/annurev.arplant.48.1.109.
- ^ Stryer, стр. 643
- ^ Stryer et al.. стр. 733–739.
- ^ а б Kuzuyama, T.; Seto H (2003). „Diversity of the biosynthesis of the isoprene units”. Natural Product Reports. 20 (2): 171—183. PMID 12735695. doi:10.1039/b109860h.
- ^ Grochowski, L.; Xu H; White, R. (2006). „Methanocaldococcus jannaschii uses a modified mevalonate pathway for biosynthesis of isopentenyl diphosphate”. Journal of Bacteriology. 188 (9): 3192—3198. PMC 1447442 . PMID 16621811. doi:10.1128/JB.188.9.3192-3198.2006.
- ^ „The”. Annual Review of Plant Physiology and Plant Molecular Biology. 50: 47—65. 1999. PMID 15012203. doi:10.1146/annurev.arplant.50.1.47.
|first1=захтева|last1=у Authors list (помоћ) - ^ а б „Sterol”. Annual Review of Biochemistry. 50: 585—621. 1981. PMID 7023367. doi:10.1146/annurev.bi.50.070181.003101.
|first1=захтева|last1=у Authors list (помоћ) - ^ Lees, N.; Skaggs B; Kirsch, D.; Bard M (1995). „Cloning of the late genes in the ergosterol biosynthetic pathway of Saccharomyces cerevisiae—a review”. Lipids. 30 (3): 221—226. PMID 7791529. doi:10.1007/BF02537824.
- ^ Stryer et al.. стр. 625–626.
Литература
[уреди | уреди извор]- Maitland, Jr Jones (1998). Organic Chemistry. W W Norton & Co Inc (Np). стр. 139. ISBN 978-0-393-97378-5.
- Međedović, S.; Maslić, E.; Hadžiselimović, R. (2002). Biologija 2. Svjetlost, Sarajevo. ISBN 978-9958-10-222-6.
- аутора, Група (2006). Угрешић, Ненад, ур. Фармакотерапијски водич 3 (PDF). Београд. ISSN 1451-4680. Архивирано из оригинала (PDF) 15. 07. 2011. г. Приступљено 23. 05. 2010.
- Кораћевић, Даринка; Бјелаковић, Гордана; Ђорђевић, Видосава. Биохемија. Савремена администрација. ISBN 978-86-387-0622-8.
- Шербан, Нада М. (2010). Биологија 1. Београд: ЗУНС. стр. 11. ISBN 978-86-17-17585-4.
- Bhagavan, NV (2002). Medical Biochemistry. San Diego: Harcourt/Academic Press. ISBN 978-0-12-095440-7.
- Devlin, T. M. (1997). Textbook of Biochemistry: With Clinical Correlations (4th изд.). Chichester: John Wiley & Sons. ISBN 978-0-471-17053-2.
- Stryer, L.; Berg, J. M.; Tymoczko, J. L. (2007). Biochemistry (6th изд.). San Francisco: W.H. Freeman. ISBN 978-0-7167-8724-2.
- van Holde KE, Mathews CK (1996). Biochemistry (2nd изд.). Menlo Park, California: Benjamin/Cummings Pub. Co. ISBN 978-0-8053-3931-4.
- Georg Löffler, Petro E. Petrides (2003). Biochemie und Pathobiochemie. Berlin: Springer. ISBN 978-3-540-42295-2.
- Florian Horn, Isabelle Moc, Nadine Schneider: Biochemie des Menschen. Thieme. . Stuttgart. 2005. ISBN 978-3-13-130883-2.
- Charles E. Mortimer, Ulrich Müller: Chemie. Thieme. . Stuttgart. 2003. ISBN 978-3-13-484308-8.
- F.D. Gunstone, J.L. Harwood, F.B. Padley: The Lipid Handbook. Chapman and Hall, London New - York. 1986. ISBN 978-0-412-24480-3.
- O.W. Thiele: Lipide, Isoprenoide mit Steroiden. G. . Stuttgart: Thieme Verlag. 1979. ISBN 978-3-13-576301-9.
- Robert C. Murphy: Mass Spectrometry of Lipids, Handbook of Lipid Research Vol. 7, Plenum Press. New York and London. 1993. ISBN 978-0-306-44361-9.
- Julian N. Kanfer and Sen-itiroh Hakomori, Sphingolipid Biochemistry, vol. 3 of Handbook of Lipid Research (1983)
- Dennis E. Vance and Jean E. Vance (eds.), Biochemistry of Lipids and Membranes (1985).
- Donald M. Small, The Physical Chemistry of Lipids, vol. 4 of Handbook of Lipid Research (1986).
- Robert B. Gennis, Biomembranes: Molecular Structure and Function (1989)
- Charles R. Scriver, Arthur L. Beaudet, William S. Sly, and David Valle, The Metabolic and Molecular Bases of Inherited Disease (1995).
- Gunstone, F. D. Fatty acids and lipid chemistry. — London: Blackie Academic and Professional, 1996. 252 pp.
- Robert M. Bell, John H. Exton, and Stephen M. Prescott (eds.), Lipid Second Messengers, vol. 8 of Handbook of Lipid Research (1996).
- Christopher K. Mathews, K.E. van Holde, and Kevin G. Ahern, Biochemistry, 3rd ed. (2000).
- Alberts, B., et al. «Essential Cell Biology, » Garland Science. (2nd изд.). 2004. ISBN 978-0-8153-3480-4..
- Solomon, Eldra P., et. al. (2005) «Biology, 7th Edition.» Thomson, Brooks/Cole.
- «Advanced Biology — Principles and Applications.» C.J. Clegg and D.G. Mackean. ISBN 978-0-7195-7670-6..
- Fahy E. et al. A comprehensive classification system for lipids // J. Lipid. Res. 2005. V. 46, № 5. P. 839—861.
Спољашње везе
[уреди | уреди извор]- ApolloLipids - Provides dyslipidemia and cardiovascular disease prevention and treatment information as well as continuing medical education programs.
- Lipids, Membranes and Vesicle Trafficking - The Virtual Library of Biochemistry and Cell Biology
- The Lipid library - provides information on the chemistry, analysis and biochemistry of lipids
- LIPID MAPS: LIPID Metabolites and Pathways Strategy
- IUPAC glossary entry for the lipid class of molecules what is IUPAC?
- "A comprehensive classification system for lipids" Архивирано на сајту Wayback Machine (24. август 2010)
- CCMDWeb, a lipid health educational resource on global risk reduction for dyslipidemia and cardiovascular disease as well as online CME programs.
- European Federation for the Science and Technology of Lipids